
par Philippe Cosentino
Pour étudier l’évolution de la fréquence allélique, il existait déjà plusieurs logiciels, la plupart hébergés sur l’académie de Nice.
Cependant, chaque logiciel avait son domaine d’application. Les divers logiciels consacrés à la dérive génétique ne permettaient pas d’envisager les effets de la sélection naturelle, et inversement, ceux prenant en compte la valeur sélective des génotypes (« Evolution allélique » par exemple) reposaient sur des modèles dont l’effectif était forcément infini.
Avec Géné’pop il est enfin possible de faire varier dans le même modèle l’effectif (qui peut être fini ou infini), les valeurs sélectives et les probabilités de mutation. On peut ainsi voir les effets combinés de ces 3 forces évolutives (les mutations, la dérive et la sélection).
Géné’pop fonctionne comme une application web, en ligne et hors-ligne (on peut télécharger l’application).
Présentation de l’interface
L’écran de Géné’pop est divisé en 3 parties.
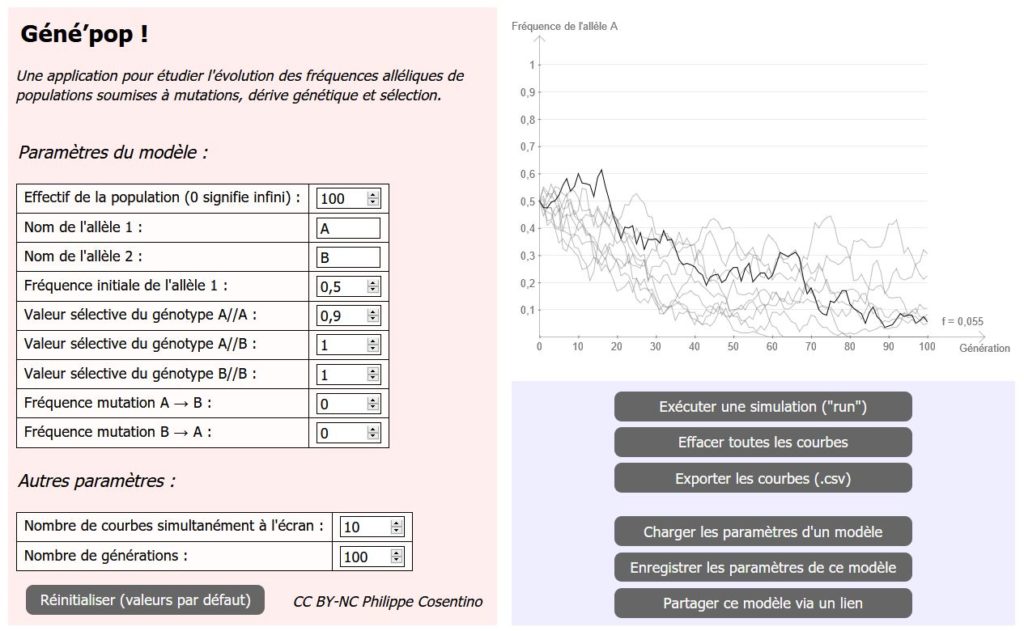
- La moitié gauche de l’écran permet de saisir tous les paramètres du modèle (l’effectif, le nom des allèles, les valeurs sélectives etc.).
- Le quart supérieur droit de l’écran correspond au graphique de suivi de la fréquence allélique.
- Le quart inférieur droit contient les boutons contrôlant le logiciel, en particulier certaines fonctions d’importation et d’exportation.
La vidéo suivante présente rapidement les possibilité de ce logiciel :
https://acamedia.ac-nice.fr/edit_video.php?vid=6406
Pistes d’exploitations
- Ecart à l’équilibre de Hardy-Weinberg
On part des réglages par défaut, et on affecte la valeur 0 à l’effectif, ce qui signifie qu’on souhaite travailler avec une population d’effectif infini.
Si on lance la simulation, on constate que la fréquence de l’allèle A reste constante au fil des générations, conformément à l’équilibre de Hardy-Weinberg.
Mais si l’on modifie un seul paramètre du modèle, on constate que la fréquence de l’allèle se met à varier.
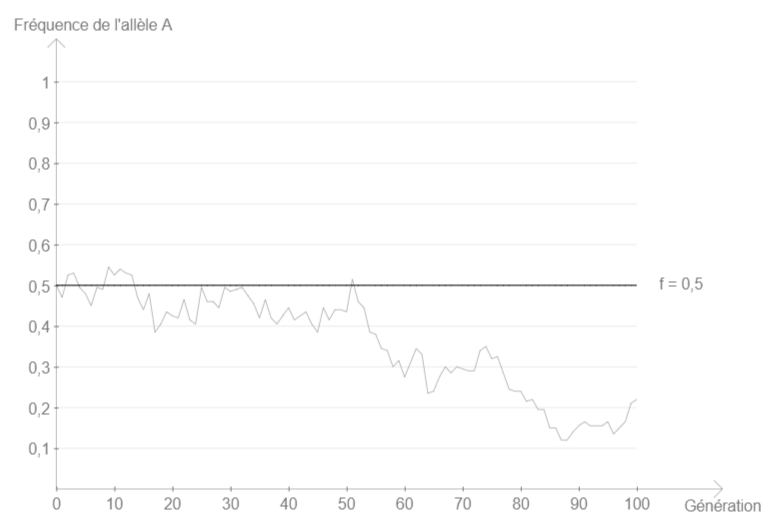
On s’écarte de l’équilibre dès lors que :
- on assigne une valeur finie à l’effectif (par exemple 50)
- on favorise un génotype par rapport aux autres (en modifiant les valeurs sélectives)
- on introduit une probabilité de mutation non nulle
- Dérive génétique
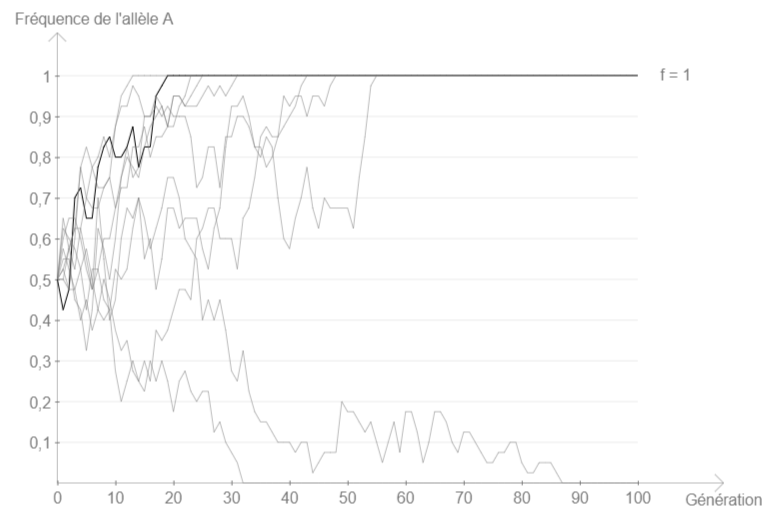
Si à partir des réglages par défaut, on lance plusieurs fois la simulation, on constate que dans certains cas, l’allèle n°1 finit par se fixer (fréquence 1) ou par disparaître (fréquence 0).
Si à présent on diminue l’effectif, par exemple en le mettant à 20, on constate que la fixation (ou la disparition) de l’allèle est bien plus rapide qu’avec 100 individus (influence de l’effectif sur la force de la dérive génétique).
Remarque : si on saisit un effectif infini (en rentrant la valeur 0), on constate que la fréquence de l’allèle ne varie pas (équilibre de Hardy-Weinberg) ; il n’y a pas de dérive.
- Sélection naturelle
Prenons l’exemple de la mucoviscidose. On saisit m et M comme noms d’allèles (ce seront deux allèles du gène CFTR, M étant l’allèle de référence).
On attribue à chaque génotype une valeur sélective différente : 1 pour M//M et M//m (phénotypes sains), et 0 pour m//m (mucoviscidose). On justifie la valeur de 0 par le fait que les individus présentant ce génotype ne peuvent pas se reproduire (valeur sélective nulle).
Pour ne pas subir la dérive génétique, on choisir un effectif infini (saisir 0).
On constate sans surprise que la fréquence de l’allèle de l’allèle m tend vers 0 au bout d’une centaine de générations.
On intègre ensuite l’hypothèse (controversée, attention), selon laquelle les individus M//m seraient en partie résistants à la fièvre typhoïde, en donnant un léger avantage au génotype M//m (valeur 1), par rapport à M//M (valeur 0,9).
On constate alors que la fréquence de l’allèle m se stabilise à une fréquence plus élevée (et surtout non nulle) que précédemment.
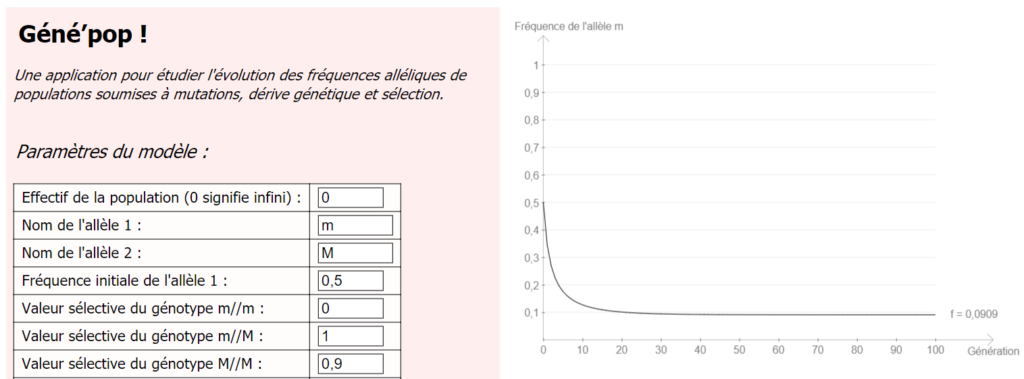
Cet avantage hétérozygote pourrait expliquer la fréquence anormalement élevée de l’allèle responsable de la mucoviscidose dans la population (l’une des maladies génétiques les plus fréquentes), mais cette résistance n’est pas prouvée.
- Pression de mutation
Même sans conférer d’avantage sélectif, les mutations peuvent à elles seules écarter une population de l’équilibre de Hardy-Weinberg.
On va considérer que l’allèle A donne une protéine fonctionnelle, tandis que l’allèle B représente tout allèle donnant une protéine non fonctionnelle.
La probabilité qu’une mutation transforme l’allèle A en allèle non fonctionnel est considérablement plus élevée que l’inverse. On considèrera pour simplifier qu’elle est négligeable (mais on peut très bien affecter une valeur non nulle).
On saisit donc une valeur de 0,01 pour la probabilité de mutation dans le sens A → B, et un effectif infini (saisir 0) pour se dédouaner de la dérive.
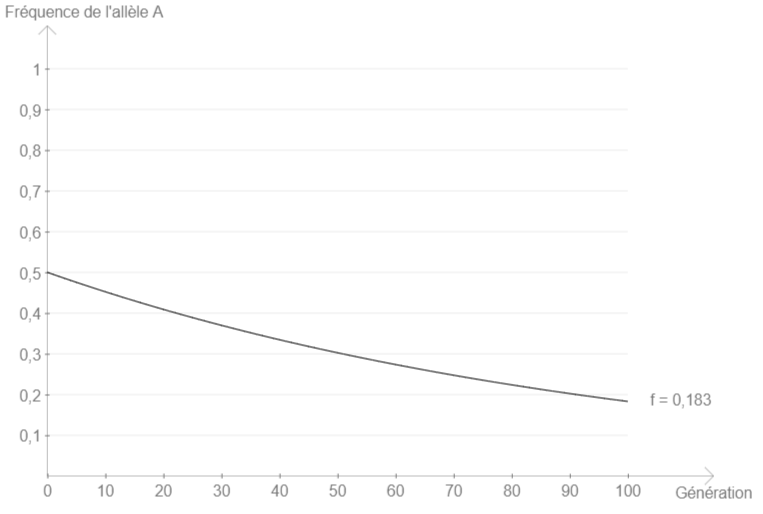
On constate alors que la fréquence de l’allèle A diminue au fil des générations (écart par rapport à l’équilibre de Hardy-Weinberg).
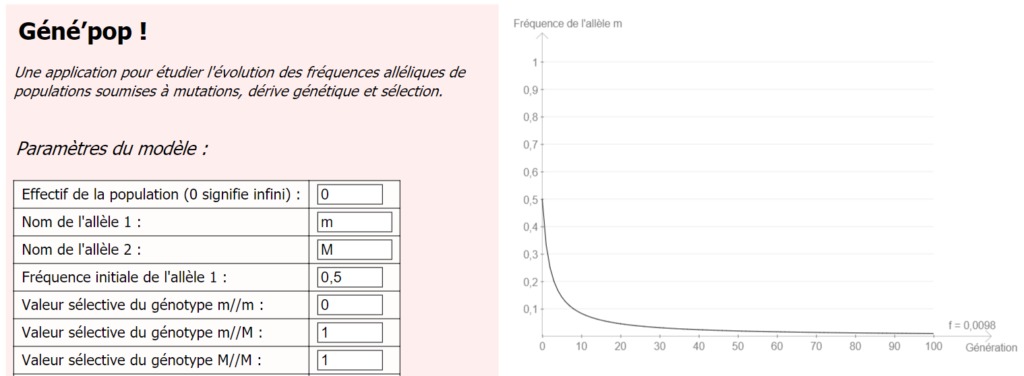
Si l’on revient à notre modèle de mucoviscidose (lien à la fin de l’article), et qu’on attribue une valeur non nulle à la probabilité de mutation de M → m, on constate que la fréquence de m ne tend plus vers 0.
Ainsi, il n’est pas nécessaire d’invoquer une quelconque « résistance des hétérozygotes » pour expliquer le fait que la fréquence de l’allèle m, responsable de la mucoviscidose, est relativement élevé (de l’ordre de 0,02 dans la population humaine réelle).
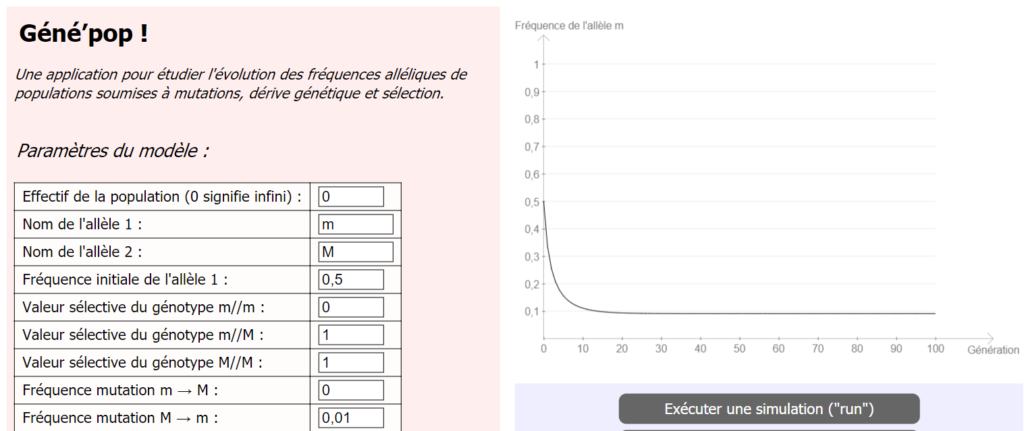
Résultats obtenus pour l’allèle m, si on prend en considération la mutation M -> m
Bilan
Il existe de nombreux logiciels équivalents à Géné’pop (aux noms similaires : popG, genPop etc.) mais généralement ils sont en anglais, utilisent Java, ou sont inabordables pour des élèves de lycée. Géné’pop n’a rien de révolutionnaire, mais rend accessible un outil de génétique des populations généraliste, « classique », à des élèves de lycée francophones.
La seule limitation de ce logiciel est qu’il ne permet de travailler que sur un couple d’allèles, et non sur 3 ou davantage comme le permet par exemple « Dérive diploïde » (qui du coup garde son intérêt intrinsèque). Il permet par contre de faire tout ce que permettait « Evolution allélique » (et bien davantage).
Il s’avèrera particulièrement utile pour étudier la sélection naturelle et la dérive génétique (surtout en classe de 2de et de terminale) mais également pour aborder différemment le modèle de Hardy-Weinberg en enseignement scientifique et en enseignement de spécialité de terminale.
Liens
Application « Géné’pop » en ligne (HTML5)
Lien direct vers le modèle « mucoviscidose » (sans avantage)
